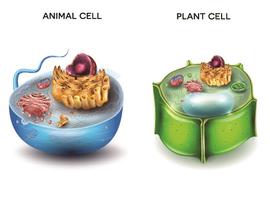
GA a lyzozomy (v rostlinách nazývané vakuoly) patří k endomembránovému systému buňky, který je dále tvořen ER a endozómy, a zahrnuje též plazmatickou membránu. Mezi složkami endomembránového systému probíhá neustálý transport a výměna, zajišťující transport a recyklaci molekul (lipidů, proteinů, syntetizovaných látek). Všechny organely spolu tedy neustále komunikují a i vznik a udržení struktury organel je na tomto transportu závislé.
Obecně lze říci, že struktura a funkce endomembránového systému jsou konzervované a lze je v obměnách nalézt ve všech eukaryotických buňkách, včetně rostlin. To se týká též Golgiho aparátu, který je důležitou organelou v živočišných i rostlinných buňkách, a hlavní rolí je modifikace a balení molekul z ER, u rostlin též syntéza polysacharidů pro stavbu buněčné stěny. Důležitým strukturním rozdílem mezi živočišným a rostlinným GA je to, že v rostlinách existuje GA jako mnohočetné nezávislé soubory cisteren, nazývané diktyosomy, které se pohybují čile cytoplazmou. Takto mobilní jednotky živočišné GA nevytváří.
Lyzozomální funkci plní v rostlinných buňkách vakuola. Jako živočišný lyzozom i rostlinné vakuoly mají jako hlavní roli degradaci proteinů, polysacharidů, lipidů či nukleových kyselin. K tomuto účelu udržují, stejně jako živočišné lyzozomy, nízké pH, hrají taktéž roli v udržování homeostáze buňky, mají zásobní funkci, podílejí se na signalizaci a autofágii. Strukturní rozdíl je ve velikosti: klasické živočišné lyzozomy jsou malé a četné struktury v buňce, rostlinné vakuoly jsou naopak obrovské organely zabírající 30-90% objemu buněk. Vznik rostlinných vakuol je vázán na fúzi membrán; ačkoliv vznik vakuol není zcela dořešen (vznikají záhy v buňkách klíčících rostlinek a narušení procesu vzniku vakuol je většinou letální), velmi pravděpodobně půjde o vznik z membrán endomembránových organel jako je ER, či post-GA struktur (endozómů, podobně jako u živočišných lyzozomů).